Kasvukohatüüpide määramise metodoloogia
Kasvukohatüüpe määratakse alati tunnuste kompleksi alusel, kus analüüsitakse nii ökotoobi (puistu kasvukoht reljeefil, muld, veerežiim), alustaimestiku kui puistu (puurinnete) omadusi ja tunnuseid. Mulla omadustest on kõige informatiivsemaks organogeense (AO, T), huumus-, leet- ja gleihorisondi esinemisel nende ulatus ja iseloom või üldse nende puudumine. Eraldise kuulumise aru- või soometsade klassi näitab turbakihi esinemine ja selle tüsedus. Eeltoodud omadustele lisaks on mulla karbonaatidesisaldus (nn kihisemise sügavus) ja lõimis. Nende kõikide alusel määratud mullaliigi järgi saab leida vastava tabeli abil võimalik kasvukohatüüp või nende grupp ja täpsustatakse see vastavalt alustaimestiku ja puistu tunnustele. Mulla morfoloogiliste tunnuste (organogeenne, huumus- ja leethorisont) tüseduse muutumist kasvukohatüüpides saab iseloomustada vastavate nivoojoontega ordinatsioonipinnal.
Metsakorralduses on ajalooliselt seistud dilemma ees: kas kasvukoha headust määratleda boniteedi või kasvutingimuste (kasvukohatüübi) järgi. Kui gradientidel tunnused muutuvad ühtlaselt-pidevalt ehk kontuitiivselt (kas sõnaliselt või arvuliselt väljendatuna), siis objekti (puistueraldise) kirjeldamisel tuleb anda hinnang üheselt, st väljavõtteliselt ühe nimetusena. Vaid reljeefi järskude muutuste korral muutuvad ka näitajad järsult (v.a tasandikel sooalade läheduses, kus vähemärgatavad seljakud sooalade sees tingivad suuri muutusi). Metsakorralduses antakse ala iseloomustus kasvukohatüübi täpsusega, teaduslikes töödes kindlasti alltüübi täpsusega, kuigi viimaseni pole sageli jõutud (hea kui tüübirühm on õieti määratud).
Puurinde parameetritest annavad tüüpide määramisel enam infot puistu boniteet, koosseis ja rinnete arv. Puistu boniteet määrab mitmel juhul teiste metsakoosluse tunnuste sarnasuse korral kuuluvuse ühte või teise kasvukohatüüpi või tüübirühma ning on esimeseks filtriks määramisel. Samal ajal näitab boniteet integreerivalt mullatingimuste muutust ja suunda eraldistel.
Juhul, kui on leitud puistute tagavarad ja vanused, siis nende järgi saab arvutada tagavara keskmise juurdekasvu. Uurimistööl suure rea kasvukohatüüpide toomisel saab tagavara keskmiste juurdekasvude võrdlemisel ka selgitada, kas vastav kasvukohatüüp on õigesti määratud. Analüütiline (arvutuslik) tegur juurdekasvu suurus on bioproduktsiooni üks näitajatest ja peegeldab mulla omadusi ehk on üks väikestest loodusseadustest. See on siis teine filter kasvukohatüübi määramisel. Tegelikkuses teaduslike artiklite algandmetest nähtub (nt süsinikuvarude arvutamisel), et sellele ei omistata mingit tähelepanu, kuigi vead kasvukohatüüpide määramisel on silmatorkavalt nähtavad.
Metsaökosüsteemi kõigi komponentide tunnuste samaaegne analüütiline kasutamine vähendab tunduvalt riski kasvukohatüüpide määratlemisel.
On diskuteeritud metsa hindamisel kasvukohatüübi määramise vajalikkuse üle, st et kas võiks piirduda vaid boniteediklassi või baaskõrgusega (kõrgus 50- või 100-aastaselt). Siin on võimalik anda kompromislik lahendus, tuues mingites kasvukoha kirjeldustes kasvukohatüübi lühendisse organogeense horisondi (O-, AT- või T-horisondi) tüseduse ja puistu boniteediklassi numbri või baaskõrguse kasvukohatüübi lühendi juurde alaindeksina.
Näiteks: Jk2-30 ja Ss100-14, kus organogeenne horisont on vastavalt 2 ja 100 cm ning baaskõrgus H100 vastavalt 30 ja 14 m.
Puuliigi tüpoloogiline informatiivsus kasvab tunduvalt, kui arvestatakse mitte ainult puistute koosseisu, vaid ka puude kasvukiirust. Männi ja kuuse keskmise kõrguse kasvukiiruse muutumist näitavad nende isoboniteedid ordinatsiooniskeemil.
Puistu liigilise koosseisu informatiivsus ja selle iseloom võivad olla väiksemad kultuurpuistutes ja peale teostatud hooldusraieid. Eestis võivad looduslike puistute enamuspuuliikideks olla harilik kuusk ja harilik mänd, aru- ja sookask, sanglepp, harilik haab, hall lepp, harilik pärn ja saar, haruharva harilik tamm, vaher ja jalakas ning künnapuu (nende levikupiirkonnad on kasvukohatüüpide ordineeritud skeemil). Nendest kliimakspuuliikideks on viljakatel muldadel harilik kuusk ja pärn, äärmustingimustes harilik mänd või sookask. Headeks kasvutingimuste indikaatoriteks on sanglepp, saar ja pärn.
Metsakasvukohatüüpide ordineeritud skeemi tsentris olevates kasvukohatüüpides, mille muld on nõrgalt happeline ja viljakas, domineerivad looduslikult segapuistud või esineb ühe kasvukohatüübi piires mitu metsatüüpi. Ordineeritud skeemi ääreosade kasvukohatüüpides, kus skeemi üla- (põhja-) osas on muld väga kuiv või all-(lõuna-) osas väga märg ning lisaks loode- ja edelaosas väga happeline ja toitainetevaene, on võimelised metsa moodustama üks või kaks puuliiki. Erandi moodustavad skeemi idaosas sinilille, naadi ja sõnajala kasvukohatüüp, kus neutraalsel, ajuti kuival (sinilille), optimaalse niiskusega (naadi) või ajuti liigniiskel (sõnajala) mullal kasvavad mitmed puuliigid hästi või väga hästi.
Normaalse arenguga muldade maatrikstabel koos seal kasvavate puistute tõenäose boniteediga vastavalt mullaliigile (R. Kõlli algskeem E. Laasi täiendustega).
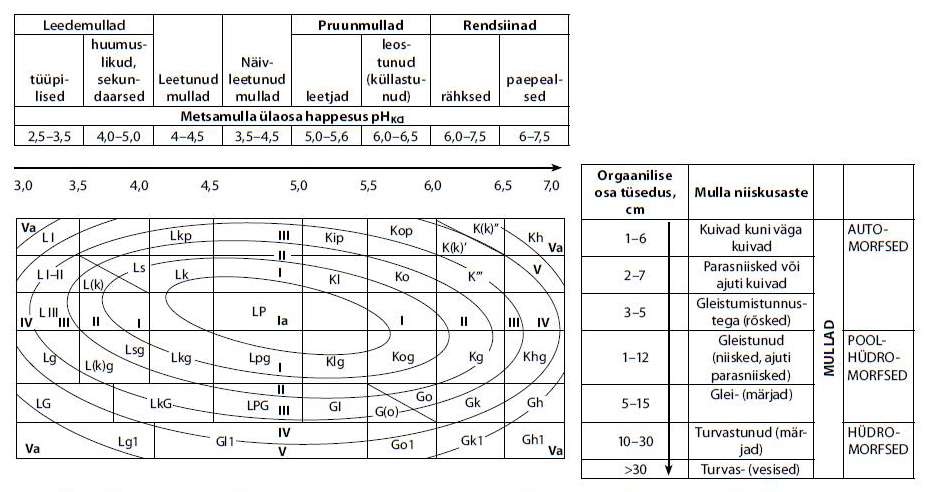
Kuna mitmele puuliigile enam-vähem sobival mullal ei ole puuliigid reastatud produktsioonikiiruse (intensiivsuse) järgi, siis soovitamegi sellistel kõrge metsakasvatusliku viljakusega muldadel (Lk, Lkg, KI, Ko, KIg, Kog, LP, LPg, kõdusoomuld) kompromissina segapuistute kasvatamist, ühtede või teistsuguste kombinatsioonidena.
Võimalikud muudatused tulevikus metsatüpoloogiliste ühikute osas
A. Katus ja E. Tappo (1965), prof V. Masing (1966), prof E. Pihelgas (1983) ning prof J. Paal (1997) on lisanud oma klassifikatsioonidesse kuivapoolsetel neutraalsetel muldadel, mis asuvad lainjatel ja künklikel aladel, veel ühe tüübirühma – sürjametsad. Viimased on mulla, alusmetsa ja alustaimestiku osas üleminekurühmaks loo-, laane- ja salumetsade vahel. Käesolevas töös on metodoloogilistel kaalutlustel võetud taas kasutusele sürjametsade tüübirühm.
Mitme metsateadlase poolt eraldatud sürjametsade tüübirühm (kuivadel kuni parasniisketel keskmise sügavusega rähkmullal ning leostunud ja leetjatel muldadel) on lubjarikastel muldadel olevatest loometsadest tootlikumate ja kuuserohkemate puistutega, viimase tõttu on sarnasus laanemetsadega. Sürjametsad tasastel aladel on tüüpilised sinilillemetsad, kõrgematel künnistel ja küngastel kuivemad sarapuumetsad. Prof E. Pihelgase (1983) esitatud klassifikatsioonis oli sürjametsade tüübirühmas sinilille ja sarapuu kasvukohatüübid.
Sürjametsad
Sürjametsad esinevad positiivsete pinnavormide – ooside ehk vallseljakute, moreenküngaste, ka voorte ja otsmoreenide – lagedel ja nõlvadel karbonaadirikkal, aga suvel läbikuivaval mullal Kesk- ja Lõuna- Eestis. Need okaspuurohked metsad kasvavad keskmise sügavusega ja sügavatel leostunud või leetjatel koreserikastel pruunmuldadel ning keskmise sügavusega rähkmullal, mis asuvad karbonaatsel liivsavi-moreenil, rähkmoreenil või kruusal, pHKCl väärtus 5,0…6,5. „Süri“ tähendab murdes kas väikest künnist, seljakut, küngast või liivast kõrgustikku. Teine tähendus on „külg“ või „serv“. Palju on kuusikuid ja männikuid ning männi-kuuse segapuistuid, vähem kaasikuid. Kuusikuid kahjustab sageli juurepess. Puistute boniteet on Ia…III, enamasti I…II. Paljud alustaimestikuliigid on lubja- ja kuiva-lembesed ning ühised loometsadega. Tüübirühma kuuluvad sinilille ja sarapuu kasvukohatüüp.
Sinilille kasvukohatüüp on paigutatud eri autoritel nii laane-, sürja- kui salumetsade tüübirühma hulka. Problemaatiline on olnud sinilille kasvukohatüübi paigutamine kas laanemetsadesse või salu-metsade tüübirühma, sest tal on palju ühiseid tunnuseid laanemetsade kõige tüüpilisema – jänesekapsa kasvukohatüübiga, sest sarnased on puistute koosseis, struktuur, tootlikkus, puuliikide vaheldumise suund (valgusnõudlike männi, arukase ja haava asendumine kuusega) ning alustaimestiku biomass ja osaliselt koosseis. Eriti sarnased on sinilille-jänesekapsa ja jänesekapsa-sinilille alltüübid, mis esinevad Otepää ja Haanja kõrgustikul. Peamine erinevus on sinilille tüübis mulla leetumise puudumine, mistõttu on mulla reaktsioon neutraalne (jänesekapsa tüübis happeline) ning sinilille kasvukohatüübi alus-taimestikus esinevad mitmed lubjalembesed taimed. Väga sage on kuuskedel kuuse-juurepessu põhjustatud juuremädanik. Seega on sinilille okaspuumetsade eripäraks kaltsifiilsemate liikidega alustaimestik, mis lähendab neid selles osas salumetsadele (sinilille-naadi alltüüp). Esineb mitmeid salutaimi: sinilill, kevadine seahernes, ussilakk, kopsurohi, naat, nurmenukk, metsaservades metsülane.
Salumetsade muldade happesus on sarnane sinilille tüübiga, kuid salumetsade koosseisus domineerivad lehtpuud ning ei esine mändi või on mänd äärmiselt juhuslik nähtus ning salumetsades pole suvel niiskuse defitsiiti, seetõttu ei tohiks olla sinilille tüübi koht salumetsade hulgas, kuna sinilillemännikuid ja männi osalusega puistuid esineb küllalt palju.
Põhjaranniku klindi jalamil kasvavad klindialused ehk pangametsad (viimast nimetust kasutas V. Masing juba 1966. a), kus J. Paal on nimetanud kuukressi kasvukohatüübi. Puurinde ja alus-taimestiku järgi on nad sarnased salumetsadele ning seega oleksid pangametsad salumetsade tüübirühma allrühmaks.
Pangametsad
Põhja-Eesti pankrannikul varieerub selle kõrgus, veerežiim, mitmesugused mullatekketingimused ja mikrokliima. Panga rusukallet ja rusuvalle katab enamasti liigirikas laialehine mets, mis kohati on kitsa ribana mitme kilomeetri ulatuses või siis katkendlikult üksikute puistutena. Pangametsad levivad Põhja-, vähem Loode-Eesti paekalda (klindi) jalami rusukaldel, kus metsariba on keskmiselt 30…60 m laiune.
Valdav osa pangaalustest muldadest kuulub rähksete rendsiinade hulka ja pangametsi iseloomustab maapinna suur kallakus, pidevalt mulla pinnale lisanduv uus lähtekivim, spetsiifiline veerežiim, kus vesi liigub horisondi siseselt kallakut mööda alla ja põhjavesi täiendab pidevalt mineraalsete toitainete varu. Mullad on suhteliselt kõrge huumusesisaldusega ja toiterikkad, veega hästi varustatud, liivsavised, neutraalsele lähedase reaktsiooniga, pHKCl = 4,5…7,5, enamikus kohtades 6,5…7,0, C : N = 9…18 (Rooma, Paal, 2001).
Pangametsad on üks väheseid enam-vähem primaarsetena säilinud metsakooslusi Eestis ja on põhjapool-seimad laialehised metsad Eestis. 1935. a eristas prof T. Lippmaa paekalda sealsete keskkonnatingimuste ja taimkatte omapära silmas pidades omaette taimegeograafiliseks alavaldkonnaks – Estonia clivosa. Esimesena käsitles pangametsi omaette metsatüübirühmana V. Masing 1966. aastal. Rakenduslikust küljest on pangametsad vähetähtsad oma väikese pindala ja majandusliku külje poolest, kuid neil on eriline koht Eesti taimestikukaardil nii geograafiliselt kui taimestiku unikaalse liigilise kompleksi poolest kõigis rinnetes.
Analüüsitabel tüübirühmade piisavuse ning kasvukohatüübi vajalikkuse ja kasutatavuse kohta
Tüübirühm
Piisavus vaid rühmast ja eripärad tüübirühma sees
Nõmmemetsad:
Täpsustamine sambliku ja kanarbiku kasvukohatüübiga ei oma erilist metsamajanduslikku tähtsust, olulisem on eristamine looduskaitselisest aspektist.
Palumetsad:
Kuna mulla viljakuses ja niiskuses on olulised erinevused pohla, jänesekapsa-pohla ja mustika kasvukohatüübi muldade osas, siis on ka uuendamistingimused ja metsakultuuride hooldamisvajadused ning tulemused erinevad.
Laanemetsad:
Kui jänesekapsa-mustika tüübi mullatingimused on kallutatud männi poole, siis jänesekapsa kasvukohatüübi mullad on kaldu kuuse ja lehtpuude poole. Kui jänesekapsa-mustika tüübis on kuusk ja mänd võrdsed ning lisanduda võib lehis, siis jänesekapsa tüübis on kuusk ja arukask võrdsed ning lisandub tamm.
Loometsad:
Loometsade kasvukohatüüpidest leesikaloo ja kastikuloo erinevad suhteliselt vähe, mullad on üksteiseks üleminevad, väliselt peegeldab olukorda (mulla niiskuse vähenemist) kõige paremini langev boniteet, millega koos uuenemistingimused järjest halvenevad. Lubikaloo tüüp erineb kahest eelnevast tunduvalt niiskema mulla poolest. Metsahoiulisest aspektist oleks vaja eristada leesikaloo tüüpi kastikuloost, sest esimeses tüübis on selgelt kaitsemetsad.
Pangametsad:
Pangametsade eraldamine kuukressi kasvukohatüübina omab vaid geobotaanilist ja loodushoiulist tähtsust.
Sürjametsad:
Sürjametsades sarapuu kasvukohatüübi eristamine praktilisel eesmärgil on raske selle väikeste pindalade tõttu, mis sageli on seotud männienamusega puistutega, omab geobotaanilist tähtsust. Sinilille tüübi suurema pindala tõttu on see sürjametsade vajalik põhitüüp. Neutraalsete muldade esinemise tõttu on metsamajanduslikult vajalik eristada (kuuse-juurepessu klassikaline riskirühm) happelise mullaga laanemetsadest.
Salumetsad:
Kui alustaimestik on sürjametsadega mõnevõrra sarnane, siis niiskusolud erinevad tunduvalt nagu ka puistute koosseis, mis on suure lehtpuude ülekaaluga. Sõnajala tüübi eristamine on raske, sest on üleminek naadi tüübilt märjema poole, peamiselt kõrge (I) boniteet, lehpuistud ja alustaimestik liidab ta salumetsade hulka.
Soovikumetsad:
Kõige raskemini eristatavate kasvukohatüüpidega tüübirühm, mis kuulub märgade gleimuldadega alade hulka, kus niiskustingimused võivad tugevasti muutuda vegetatsiooniperioodi jooksul. Eelneva tüübirühma sõnajala ning soovikumetsade angervaksa, tarna-angervaksa, tarna ja osja tüüpide vahel on olulisteks erinevusteks soovikumetsade puistute madalam boniteet ja ka mändide märgatavam esinemine soovikumetsades. Esinevad peamiselt leht-okaspuu- ja lehtpuu-segametsad.
Rabastuvad metsad ehk kõrvemetsad:
Madalaboniteedilised märja happelise mineraalmullaga männikud. See tüübirühm asub nõmme- ja palumetsade ning teiselt poolt samblasoometsade vahel, kus ühenduslüliks palumetsadega on karusambla-mustika tüüp (analüütiliselt peaks leede-gleimullal asuma uus formatsioom – sookailu tüüp). Rühma pika nime asemele sobiks nimetus „kõrvemetsad“.
Lammimetsad:
Vaid suuremate üleujutustega jõgede lammidel esinevad metsad. Sarnasus on lodu ja sõnajala tüübiga. Tähtsus vaid geobotaanilisest ja klassifitseerimise aspektist.
Rohusoometsad:
Lehtpuude domineerimisega rohttaimestikurikaste ja hästi lagunenud turbaga rohusoometsadeeristamine samblasoometsadest on vajalik.
Samblasoometsad:
Männienamusega puhmasterohkete halvasti lagunenud turbamullaga soometsade tüübirühm on vajalik nii geobotaanilisest, metsatüpoloogilise klassifitseerimise kui loodushariduse aspektist.
Kõdusoometsad:
Metsamajanduslikult oluline eristada hästilagunenud turbakihiga tootlikuimad soometsad samailmelistest kõrgeboniteedilistest mineraalmullal asuvatest tüübirühmadest.
Kui minna lihtsustamise teed ja kasutada vaid tüübirühmasid (11 on tavatüübirühmi pluss panga- ja lammimetsad), siis saab ka asjad aetud ja pilt kasvukohatingimustest on enam-vähem olemas. Kõige suuremaks probleemiks on palumetsade rühm, kus vaatamata ühisele peapuuliigile (mänd) ja leede-muldadele on küllalt suured erinevused mulla niiskustingimustes.
Metsatüpoloogia ühikud kaheks erinevaks otstarbeks
| Metsamajndamise tarbeks | Õppe-teaduslik tüpogeeniline süsteem | ||
| Tüübirühmad | Kasvukohatüüp | Tüübirühmad | Kasvukohatüüp |
| Nõmmemetsad | Nõmme | Nõmmemetsad | Sambliku |
| Kanarbiku | |||
| Palumetsad | Pohla | Palumetsad | Pohla |
| Kilpjala | Kilpjala (jänesekapsa-pohla) | ||
| Mustika | |||
| Laanemetsad | Leselehe | Laanemetsad | Leselehe (Jänesekapsa-mustika) |
| Jänesekapsa | Jänesekapsa | ||
| Loometsad | Loo | Loometsad | Leesikaloo |
| Kastikuloo | |||
| Lubikaloo | |||
| Pangametsad | Kuukressi | ||
| Sürjametsad | Sinilille | Sürjametsad | Sinilille |
| Sarapuu | |||
| Salumetsad | Naadi | Salumetsad | Naadi |
| Sõnajala | Sõnajala | ||
| Soovikumetsad | Angervaksa | Soovikumetsad | Angervaksa |
| Tarna-osja | Tarna | ||
| Tedremarana (osja) | |||
| Kõrve- ehk rabastuvad metsad | Sinika | Kõrve- ehk rabastuvad metsad | Sookailu (karusambla mustika, mustika sinika+ sinika mustika) |
| Karusambla | Karusambla | ||
| Sinika | |||
| Lammimetsad | Humala | ||
| Pika tarna (lodu) | |||
| Rohusoometsad | Lodu | Rohusoometsad | Lodu |
| Madalsoo | Madalsoo | ||
| Samblasoometsad | Siirdesoo | Samblasoometsad | Siirdesoo |
| Raba | Raba | ||
| Kõdusoometsad | Mustika kõdusoo | Kõdusoometsad | Mustika kõdusoo |
| Jänesekapsa kõdusoo | Jänesekapsa kõdusoo | ||
| Kokku 11 | 20 | 13 | 29 |